|
Heslop-Harrison Group Homepage Science Bovid (cow) diversity People Methods/techniques In situ Hybridization Molecular methods- lab protocols Technology development Courses and Science Pictures Journal editing Policy Information/Publications Jack Heslop-Harrison Links Site information
|
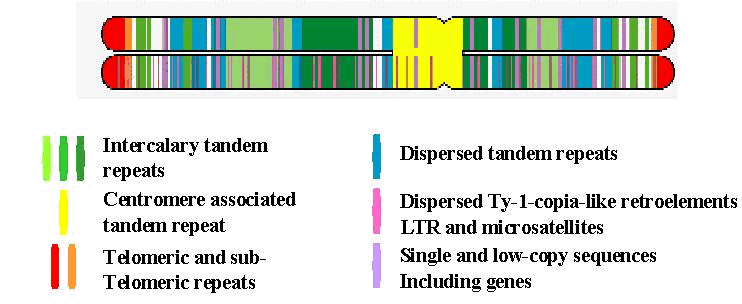
The Linear Chromosome ModelThomas Schmidt (University of Dresden from 4/2003) and Pat Heslop-Harrison Plants over wide taxonomic groupings have similar genes and ordering of genes along the chromosomes. However, the repetitive DNA, much of no known function and often constituting the majority of the genome, varies extensively from species to species in absolute amount, sequence and dispersion pattern. Despite this, it is known that families of repeated DNA motifs each have a characteristic genomic location within a genus, and that there are different constraints on the evolution of repetitive DNA and genes. There is now enough data about different repetitive DNA - from sequencing, Southern analysis and in situ hybridization - to build a model of the organization of a typical plant genome, which is useful for gene cloning, evolutionary studies and gene transfer. Sugar beet is a valuable model species for investigating the large-scale chromosomal and molecular organization of the nuclear genome, because it is diploid (2n=18, where n is the basic chromosome number of the haploid genome), a relatively small genome (750 Mbp) that is representative of many genomes. The large number of families of repetitive DNA, their high amplification and different characteristic locations fill most of the chromosome with repetitive DNA. Together, the data from in situ hybridizations and molecular analyses suggest the integrated model for plant chromosome organization presented below.
Different classes of repetitive DNA have characteristic genomic distributions. Genes are clustered in discrete blocks between the various repetitive DNA motifs, each of which has a characteristic location and genomic organization. Ty1-copia and other families of long terminal repeat (LTR) retroelements may comprise 50% of the genome. The LTR retroelements are located throughout the genome with some clusters and depleted regions, perhaps a consequence of different evolutionary rates of sequence amplification or targeting of insertion sites. Simple sequence repeats or microsatellites are dispersed and present as clusters (shown only on the lower chromatid and varying between motifs), as well as occurring within arrays of larger tandem repeats. The 18S-5.8S-25S and 5S rRNA genes are clustered on one or more chromosome pairs in the genome (not drawn). Most plant chromosomes have the 7 bp telomeric nucleotide sequence repeated at their ends. Within a plant genome, all chromosomes have a rather similar composition and organization of repeats because of homogenization of the genome, although copy numbers of individual repeat motifs may vary between chromosomes. In sugar beet, where the average chromosome has about 80 Mbp of DNA, all the genomic elements illustrated have been found, and the species provides the reference upon which the model is based. Wheat, rye and barley, with chromosomes ten times larger, follow the same model: families of tandem repeats and the other sequence classes with characteristic locations have been found. In plants with small genomes, such as Arabidopsis (chromosome size 15 to 30 Mbp), major intercalary tandem repeats are not known; tandemly repeated DNA is clustered around the centromere and retrotransposons are more dispersed.
|