| Heterochrony in cavusgnathid conodonts | ||||||||||
 |
are the "Granton Animals" paedomorphic?
Mark A. Purnell |
|||||||||
 Recent reconstructions of the conodont body, all approximately
4 cm long. a, from Aldridge and Purnell3; b, from Discover5; c, from Purnell et al.6; d, from Purnell7; e, from Aldridge et al.2 Recent reconstructions of the conodont body, all approximately
4 cm long. a, from Aldridge and Purnell3; b, from Discover5; c, from Purnell et al.6; d, from Purnell7; e, from Aldridge et al.2 |
||||||||||
| The discovery and interpretation of the Scottish fossils preserving traces of conodont soft tissues1, 2 have revolutionized many aspects of the study of conodonts. After more than 100 years of uncertainty and speculation, we finally have some understanding of the biology and ecology of conodonts. Reconstructions of the conodont body are an important component of this new conceptual framework. Not only do these reconstructions satisfy our basic curiosity about a group of enigmatic, extinct organisms, they are a powerful influence on developing concepts of conodonts as living animals, and reconstructions themselves become the subject of biological interpretation and speculation. | ||||||||||
|
||||||||||
| Almost all recent reconstructions of conodonts (see above) are based primarily on the best known and best preserved material: the Carboniferous cavusgnathids (Clydagnathus windsorensis) from Granton, Scotland. But how well do these few specimens represent conodonts as a whole? | ||||||||||
 |
||||||||||
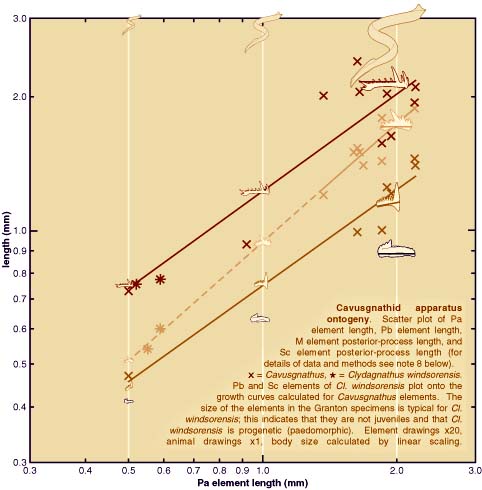
| Analysis of apparatus growth in cavusgnathid conodonts (see plot above) using measurements of elements in the Granton specimens and bedding plane assemblages from the Carboniferous of Montana (see photographic images above plot), provides strong support for the hypothesis that Clydagnathus windsorensis is progenetic (i.e. paedomorphic). This does not call into question any of the key anatomical attributes of conodonts, or their assignment to the vertebrates3. Neither does it indicate that the Granton conodonts are larval forms (contra ref. 4). It does suggest, however, that some details and proportions of reconstructed animals may reflect juvenile morphology rather than that of a typical adult conodont, whatever that may be (see plot). | ||||||||||
| REFERENCES AND NOTES 1. Briggs, D. E. G., Clarkson, E. N. K., and Aldridge, R. J., 1983. The conodont animal. Lethaia, 16, 1-14. 2. Aldridge, R. J., Briggs, D. E. G., Smith, M. P., Clarkson, E. N. K., and Clark, N. D. L., 1993. The anatomy of conodonts. Philosophical Transactions of the Royal Society of London, Series B, 340, 405-421. 3. Aldridge, R. J., and Purnell, M. A., 1996. The conodont controversies. Trends in Ecology and Evolution, 11, 463-468. 4. Forey, P., and Janvier, P., 1994. Evolution of the early vertebrates. American Scientist, 82, 554-565. 5. Picture blood on its teeth. Discover, 17, 45 (1996). 6. Purnell, M. A., Aldridge, R. J., Donoghue, P. C. J. & Gabbott, S. E. 1995: Conodonts and the first vertebrates. Endeavour 19, 20-27. 7. Purnell, M. A. 1995: Large eyes and vision in conodonts. Lethaia 28, 187-188. 8. Element dimensions are taken from bedding plane assemblages of Cavusgnathus from the middle Carboniferous of Montana, USA, and from exceptionally preserved Clydagnathus windsorensis from the Lower Carboniferous of Scotland. Curves represent the power function or allometric equation y=axb and were fitted to Cavusgnathus data only, using linear regression. Dashed line extrapolates the Pb element curve beyond the Cavusgnathus data. |
||||||||||


